
Winemaking involves the multiplication and metabolism of numerous yeasts and bacteria in grape juice. After fermentation, the fermented juice is stabilized by the addition of sulfites (sulfur dioxide) to protect against refermentation or the growth of deleterious microorganisms that could spoil the wine through production of undesirable flavors and aromas.[1] Sulfur dioxide (SO2) is an antioxidant and antimicrobial,[2] and it is also used to sanitize and store empty barrels. SO2 has not been systematically evaluated to sanitize wine cooperage, and current knowledge of the use of SO2 is largely based upon studies performed in wine, where carbonyl compounds, sugars and other uncontrolled conditions may confound its observed antimicrobial effects. Glucose present in wine is one of the main SO2-binding compounds. Moreover, SO2 forms additional compounds with aldehydes and, to a lesser extent, with ketones.[3] SO2 exists in equilibrium in aqueous solutions between molecular SO2 (SO2•H2O), bisulfite (HSO3-) and sulfite (SO2-3) species, but this equilibrium is strictly dependent on pH.[2,4] It is generally believed that the molecular sulfur species is the antimicrobial form of SO2, which is predominant at low pH. Because SO2•H2O does not have a charge, the molecule enters the cell and undergoes rapid pH-driven dissociation at cytoplasmic pH (generally near 6.5) to yield bisulfite and sulfite. As the intracellular concentration of molecular SO2 decreases due to the internal equilibrium, more molecular SO2 enters the cell, further increasing intracellular concentrations.[4]
Oak barrels have been commonly used for aging wines and spirits due to their positive effects on the finished product that include increased color stability, spontaneous clarification and desirable complex aromas.[5] However, spoilage sometimes occurs during aging, or even after bottling, due to the continued growth or residual secondary metabolites of contaminant microorganisms, such as the yeast Dekkera/Brettanomyces, causing widespread losses in the wine industry due to degraded wine quality.[1] The growth of yeasts belonging to Dekkera⁄Brettanomyces during the production of red wine, especially during aging, can seriously affect the organoleptic quality of the finished product. These yeasts grow slowly during wine aging in wooden barrels, particularly when the SO2 concentration is low (molecular SO2 < 0.5 mg/L), the pH is high (> 3.8) and the temperature is above 15 °C.[6] Although barrel disinfection methods are becoming more sophisticated (water vapor, ozonization, etc.), the shape and microstructure of wooden barrels offer undesirable microorganisms niches that provide a great degree of protection from direct contact by sanitizers.[7] It is now generally accepted that control of Dekkera/Brettanomyces spp. cannot be achieved by mere cleaning or incomplete sanitization of all cellar equipment but demands much more stringent microbiological control and judicious utilization of sulfite or dimethyl dicarbonate.[8] The ability of yeast strains to survive high alcohol concentrations allows them to carry out refermentation in barrels or bottles, thereby modifying the alcohol/sugar balance and the aroma of the wine, reducing its quality. This process is mainly carried out by Zygosaccharomyces bailii and Saccharomyces cerevisiae, due to their tolerance of high concentrations of alcohol and SO2[9] and their survival of primary alcoholic fermentation.[10] The presence of yeasts of the genus Zygosaccharomyces, in particular Z. bailii, is well known in wineries producing sweet or sparkling wines using juice concentrate or sulfited grape juice. Their high level of resistance to preservatives, particularly in Z. bailii, means that addition of high, but sublethal, doses decreases the incidence of competing microorganisms, making the survival of resistant strains even more problematic.[8] To reveal more precise indications of the true antimicrobial efficacy of SO2, we conducted in vitro experiments in which no other compound was added other than SO2 in the form of potassium metabisulfite (KMB). SO2 was also evaluated using 5-g sulfur discs in wine barrels (in vivo experiments). The purpose of the study was to test SO2 at three pH levels to assess its efficacy against wine-spoilage yeasts and then assess the use of sulfur discs (~5 g) in naturally contaminated barrels held for 3 and 6 weeks. The results from these experiments will serve as a guide to more efficient sanitization practices using SO2 as a sanitizer.
Methods
Microorganisms
Three isolates of Dekkera/Brettanomyces bruxellensis (2080, CE149 and CE261), three isolates of S. cerevisiae (CE78, CE9 and CE81) and an isolate of Z. bailii (4A1) were obtained from the Department of Food Science collection at Cornell University.
Preparation of starter culture and inoculation
The yeasts, stored at -80 °C in 15% glycerol (w/v), were revitalized and maintained on YPD agar. All strains were grown until stationary phase (108 CFU/mL) or at least 106 CFU/mL (growth under agitation at 200 rpm, 30 °C). The growth time varied according to the strain. Each strain was grown in YPD broth adjusted to different pH levels (3.0, 3.2 and 3.4) with 1 M HCl and/or 1 M NaOH. Once the cultures reached 108 or 106 CFU/mL, the target inoculum was verified via a viability assessment. A 1-mL volume of culture was placed in sterile Eppendorf tubes and centrifuged (4,500 rpm, room temperature); the supernatant was discarded and resuspended in 1 mL McIlvaine buffer[11] at the different pH levels stated above. A 1-mL culture was inoculated into 99 mL McIlvaine buffer at the different pH levels mentioned above, reaching a concentration of 104–106 CFU/mL. Analyses were performed in triplicate.
Preparation of KMB solution
The solutions were prepared before each experiment by dissolving the required amount of KMB in McIlvaine buffer at the different pH values. The final concentrations of KMB were 0 and 300 mg/L. Each KMB solution was spiked with 1 mL cells to achieve a final volume of 100 mL and plugged with rubber stoppers.
Sampling and microbiological isolation
Each flask was aseptically sampled at different times. The flasks were incubated in a water bath at 30 °C with no agitation and sampled at 0, 15, 30, 60 and 90 minutes. At each sampling time, 1 mL was taken and properly diluted in 0.1% (w/v) buffered peptone water, immediately plated in duplicate onto YPD agar and incubated at 30 °C for 48–72 hours for Z. bailii and S. cerevisiae, and up to 3–4 weeks for D./B. bruxellensis. When necessary, direct plating from the flask was also performed to increase the level-of-detection threshold. The volume plated was 100 µL.
Microbiological enumeration
Plates were enumerated for total microbial count. The counts were averaged and expressed as log values. The log reduction was then calculated for each strain and expressed as log values. Each inactivation experiment and controls were performed in triplicate for each strain.
Sulfur disc treatment of barrels
The 20 donated barrels used for this study were identified with Dekkera/Brettanomyces by the wineries that donated them. These naturally contaminated barrels were split in two groups of 10 barrels each to identify both Brettanomyces and general yeast populations and were then treated with sulfur discs. Each barrel had an identification number. The treatments were conducted for 3 and 6 weeks. Briefly, 7 L distilled water were added to each barrel, before and after burning the sulfur discs inside the barrels. The barrels were rolled several times to enhance the contact of water with the inner surface of the barrel, stored bung side up for 24 hours and then sampled before and after burning the sulfur discs. Water samples were collected in sterile bottles. The first portion of the water was discarded to “rinse” the bunghole (outer portion of the bung), and then 70% ethanol was sprayed around the bunghole. The samples were placed at 4 °C until analysis. The samples were analyzed to determine the initial and final yeast populations, either by filtration using discs or pertinent dilutions of the samples, since the microbial loads varied for each barrel. If samples needed to be diluted, 0.1% (w/v) buffered peptone water was used.
The samples were divided into two groups and filtered through cellulose. The results of the two filters were then averaged. The maximum volume filtered was 100 mL, and the results were calculated as CFU/100 mL and then transformed into log10. The cellulose filters were aseptically placed onto WL and YPD agars. WL agar was used to detect D./B. bruxellensis and incubated at 30 °C for up to 3–4 weeks; colonies that grew before 3 days were discarded. Incubation time was also used to demonstrate growth, as nothing that grows before 3 days in WL with cycloheximide is D./B. bruxellensis. The WL agar contained 10 mg/L cycloheximide to allow for selection of D./B. bruxellensis (dissolved in 50% ethanol and filter-sterilized), 150 mg/L biphenyl to prevent the growth of mold, 30 mg/L chloramphenicol to prevent the growth of lactic acid bacteria and 25 mg/L kanamycin sulfate to inhibit the growth of acetic acid bacteria. YPD agar was used to detect general yeast populations and was incubated at 30 °C for 48–72 hours. The YPD agar contained all of the above selective agents except cycloheximide.
Statistical analysis
The reductions in yeast were calculated from the initial concentration of yeast cells (target inoculum) at time zero minus the last concentration of yeast at time 90 minutes for the in vitro experiments. All experiments, including the controls, were performed in triplicate. The analysis used was a three-way analysis of variance, and simple comparisons were performed using the Tukey’s test (P < 0.05). For reduction of Brettanomyces and general yeast populations using sulfur discs, a Fisher’s exact test was performed.
Results and Discussion
Effects of KMB on strains
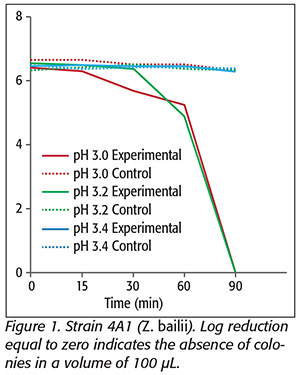 Sulfite acts as a powerful antimicrobial agent and is highly toxic to most non-Saccharomyces yeasts.[12] In this work, we studied the effects of pH at a high concentration of KMB using a matrix with no SO2-binding compounds commonly found in wines. At pHs 3.0 and 3.2, total log reductions were observed for Z. bailii. However, at pH 3.4, there was no reduction, likely due to the decreased availability of SO2, which is the germicidal form of the sanitizer (Figure 1). Divol et al.[13] found that 200 mg/L SO2 did not affect the capacity of Z. bailii cells to grow on a solid medium, but did not mention pH as a factor, which has an important influence on the antimicrobial effect of SO2.
Sulfite acts as a powerful antimicrobial agent and is highly toxic to most non-Saccharomyces yeasts.[12] In this work, we studied the effects of pH at a high concentration of KMB using a matrix with no SO2-binding compounds commonly found in wines. At pHs 3.0 and 3.2, total log reductions were observed for Z. bailii. However, at pH 3.4, there was no reduction, likely due to the decreased availability of SO2, which is the germicidal form of the sanitizer (Figure 1). Divol et al.[13] found that 200 mg/L SO2 did not affect the capacity of Z. bailii cells to grow on a solid medium, but did not mention pH as a factor, which has an important influence on the antimicrobial effect of SO2.
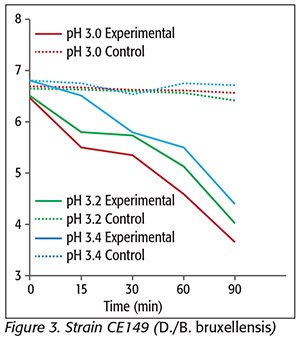
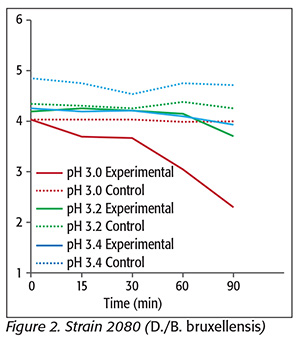 For strain 2080 (D./B. bruxellensis) (Figure 2), a 1.9-log reduction was achieved at pH 3.0. At pHs 3.2 and 3.4, there were log reductions of only 0.5 and 0.3, respectively. With regard to strain CE149 (D./B. bruxellensis) (Figure 3), there was a 3-log reduction at pH 3.0, and 2.5- and 2.4-log reductions at pHs 3.2 and 3.4, respectively. CE261 (D./B. bruxellensis) (Figure 4) was reduced 1.78 log units at pH 3.0, and 1.2 and 1.0 logs at pHs 3.2 and 3.4, respectively. Interestingly, reports about the effects of SO2 on D./B. bruxellensis inactivation are often contradictory. Some authors refer its sensitivity to values higher than 30 mg/L SO2, while others state it should be regarded as resistant and growth has been reported when values greater than 30 mg/L SO2 are used. This controversy probably arises from differences in experimental conditions and strain behavior variability.[14] Our results showed that 300 mg/L KMB was not sufficient to reduce the yeasts tested to undetectable levels.
For strain 2080 (D./B. bruxellensis) (Figure 2), a 1.9-log reduction was achieved at pH 3.0. At pHs 3.2 and 3.4, there were log reductions of only 0.5 and 0.3, respectively. With regard to strain CE149 (D./B. bruxellensis) (Figure 3), there was a 3-log reduction at pH 3.0, and 2.5- and 2.4-log reductions at pHs 3.2 and 3.4, respectively. CE261 (D./B. bruxellensis) (Figure 4) was reduced 1.78 log units at pH 3.0, and 1.2 and 1.0 logs at pHs 3.2 and 3.4, respectively. Interestingly, reports about the effects of SO2 on D./B. bruxellensis inactivation are often contradictory. Some authors refer its sensitivity to values higher than 30 mg/L SO2, while others state it should be regarded as resistant and growth has been reported when values greater than 30 mg/L SO2 are used. This controversy probably arises from differences in experimental conditions and strain behavior variability.[14] Our results showed that 300 mg/L KMB was not sufficient to reduce the yeasts tested to undetectable levels.
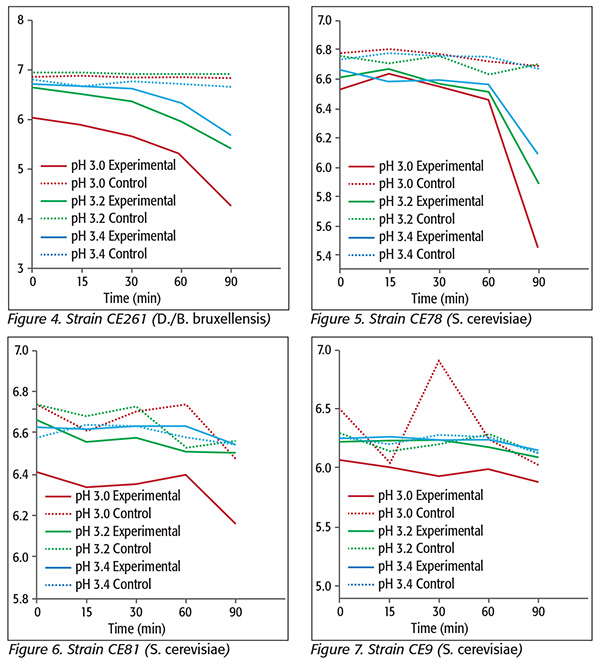
 At pH 3.0, only strain CE78 (S. cerevisiae) was reduced 1.0 log10 unit, and at pHs 3.2 and 3.4, there were reductions of only 0.7 and 0.5 log10 units, respectively (Figure 5). The two S. cerevisiae strains CE81 and CE9 were reduced only 0.2 and 0.1 log10 units, respectively, at pH 3.0 (Figures 6 and 7). No significant differences were found for either strain when three different pHs were compared using either 0 or 300 mg/L KMB, since we observed that KMB did not cause noticeable reductions in either strain. In fact, the effect of the sanitizer was expected to be negligible for S. cerevisiae strains. In reality, S. cerevisiae strains could be considered as controls since they are generally recognized as being sulfite-resistant yeasts. For S. cerevisiae, differences in resistance have been attributed to production of compounds, particularly acetaldehyde, that bind sulfite to form α-hydroxysulfonates.15 Indeed, S. cerevisiae strains produce relatively high levels of acetaldehyde.[16] However, the study of production of sulfite-binding compounds is beyond the scope of this study.
At pH 3.0, only strain CE78 (S. cerevisiae) was reduced 1.0 log10 unit, and at pHs 3.2 and 3.4, there were reductions of only 0.7 and 0.5 log10 units, respectively (Figure 5). The two S. cerevisiae strains CE81 and CE9 were reduced only 0.2 and 0.1 log10 units, respectively, at pH 3.0 (Figures 6 and 7). No significant differences were found for either strain when three different pHs were compared using either 0 or 300 mg/L KMB, since we observed that KMB did not cause noticeable reductions in either strain. In fact, the effect of the sanitizer was expected to be negligible for S. cerevisiae strains. In reality, S. cerevisiae strains could be considered as controls since they are generally recognized as being sulfite-resistant yeasts. For S. cerevisiae, differences in resistance have been attributed to production of compounds, particularly acetaldehyde, that bind sulfite to form α-hydroxysulfonates.15 Indeed, S. cerevisiae strains produce relatively high levels of acetaldehyde.[16] However, the study of production of sulfite-binding compounds is beyond the scope of this study.
Reduction of Brettanomyces and general yeast populations using sulfur discs
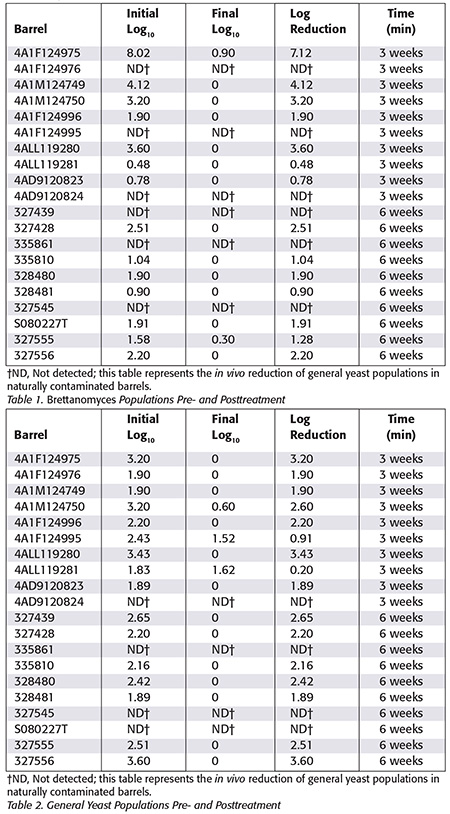
The 6-week treatments were shown to be highly effective in decreasing both Brettanomyces and general yeast populations (Tables 1 and 2, respectively) with the exception of only one barrel that presented reminiscent yeast populations after the application of sulfur discs. Three barrels of general yeast populations from the 3-week treatment had reminiscent populations of yeast after treatment. The reminiscent populations of yeasts in some barrels after treatment with sulfur discs can be attributed to an incomplete burning of the sulfur discs, since data showed that there were barrels with higher initial microbial loads that had total elimination or undetectable levels of microorganisms, and others that had fewer microorganisms and did not have a total elimination. No statistical differences were found between the 3- and 6-week treatments for both Brettanomyces and general yeast populations, suggesting that the level of disinfection was the same. SO2 rings used and held for 3 or 6 weeks were sufficient to decrease the microbial loads to undetectable levels in the majority of the barrels. However, several variables can influence treatment effectiveness when microbial elimination is taken into consideration, that is, the amount of oxygen consumed inside the barrel, the initial microbial load, the presence of debris or residues of organic matter and how much of the sulfur disc was burned.
Conclusions
In this study, we considered the use of a noninterfering matrix to be an important variable to prove the maximum efficacy of SO2 whether in solution or in the headspace of a 225-L barrel. The highly variable results found in the literature make it difficult to precisely define an optimal protocol for the use of SO2 as a surface sanitizer in wineries. When used in solution, SO2 provides relative efficacy toward the strains and initial microbial concentration levels presented here. This fact is of substantial importance for winemakers when SO2 is to be used in the liquid form at a specific pH. However, under laboratory conditions, SO2 was under no other conditions other than the ones encountered in the barrel. This should be considered when sanitization protocols are going to be achieved. The lack of a systematic approach to evaluating SO2 in common protocols of winery sanitization has resulted in the generalized use of certain protocols that might not be applicable to all the surfaces in wineries. High microbial loads were used under laboratory conditions, representing the worst-case scenario of barrel contamination. However, when we extrapolated this same approach under winery conditions, we realized that the use of SO2 as a gas, although effective, was only challenged with up to 103 CFU/mL for general yeast populations and up to 108 CFU/mL for Brettanomyces populations. This should be taken into consideration by winemakers since microbial loads in naturally contaminated barrels may vary according to their previous sanitization practices.
Winemakers must keep in mind that prevention is more effective than correction. Presanitization practices (rinsing at the right temperatures to avoid thermally adhering debris on the surface) and the correct concentration of SO2 that is effective for the surface to be sanitized are the best approaches when optimal sanitization is desired. In general, winemakers and the wine industry should consider reassessing their practices in terms of the use of scientifically validated information and utilize it to protect their wine from future contamination.
This work was supported by CONACYT-Mexico, federal formula multi-state project under grant NC-1023 and Cornell University Research Travel Awards. The authors thank the industry donors at Constellation Brands, California. Some experimental details were omitted for brevity; please contact the authors for more information.
Aguilar Solis Maria de Lourdes Alejandra, Ph.D., is a technical compliance specialist who works in the wine industry.
David M. Gadoury, Ph.D., is a senior research associate in the Department of Plant Pathology and Plant-Microbe Biology, Cornell University, New York State Agricultural Experiment Station.
Randy W. Worobo, Ph.D., is an associate professor of food science and microbiologist in the Department of Food Science, Cornell University, New York State Agricultural Experiment Station.
References
1. Lustrato, G et al. 2010. “Inactivation of Wine Spoilage Yeasts Dekkera bruxellensis Using Low Electric Current Treatment (LEC).” J Appl Microbiol 109:594–604.
2. Usseglio Tomasset, L. 1992. “Properties and Use of Sulphur Dioxide.” Food Addit Contam 9:399–404.
3. King, AD Jr. et al. 1981. “Factors Affecting Death of Yeast by Sulfur Dioxide.” J Food Prot 44:92–97.
4. Fugelsang, KC and CG Edwards. 2007. “Managing Microbial Growth,” in Wine Microbiology: Practical Applications and Procedures, eds. Fugelsang, KC and CG Edwards (New York: Springer Science Business Media), 66–67.
5. Rodriguez Rodriguez, P and E Gomez Plaza. 2011. “Effect of Volume and Toast Level of French Oak Barrels (Quercus petraea L.) on Cabernet Sauvignon Wine Characteristics.” Am J Enol Viticult 62:359–365.
6. Benito, S et al. 2009. “A Method for Estimating Dekkera⁄Brettanomyces Populations in Wines.” J Appl Microbiol 106:1743–1751.
7. Suarez, R et al. 2007. “The Production of Ethylphenols in Wine by Yeasts of the Genera Brettanomyces and Dekkera: A Review.” Food Chem 102:10–21.
8. Loureiro, V and M Malfeito-Ferreira. 2003. “Spoilage Yeasts in the Wine Industry.” Int J Food Microbiol 86:23–50.
9. Salinas, F et al. 2009. “Taqman Real-Time PCR for the Detection and Enumeration of Saccharomyces cerevisiae in Wine.” Food Microbiol 26:328–33.
10. Divol, B et al. 2006. “Genetic Characterization of Strains of Saccharomyces cerevisiae Responsible for Refermentation in Botrytis-Affected Wines.” J Appl Microbiol 100:516–526.
11. Dawson, RMC et al. 1986. Data for Biochemical Research (Oxford, UK: Oxford Science Publications).
12. Mendoza, LM et al. 2009. “Influence of Wine-Related Physicochemical Factors on the Growth and Metabolism of Non-Saccharomyces and Saccharomyces Yeasts in Mixed Culture.” J Indust Microbiol Biotechnol 36:229–237.
13. Divol, B et al. 2005. “Effectiveness of Dimethyldicarbonate to Stop Alcoholic Fermentation in Wine.” Food Microbiol 22:169–178.
14. Barata, A et al. 2008. “Survival Patterns of Dekkera bruxellensis in Wines and Inhibitory Effect of Sulfur Dioxide.” Int J Food Microbiol 121:201–207.
15. Pilkington, BJ and AH Rose. 1988. “Reactions of Saccharomyces cerevisiae and Zygosaccharomyces bailii to Sulphite.” J Gen Microbiol 134:2823–2830.
16. Romano, P et al. 1994. “Acetaldehyde Production in Saccharomyces cerevisiae Wine Yeasts.” FEMS Microbiol Lett 118:213–218.



